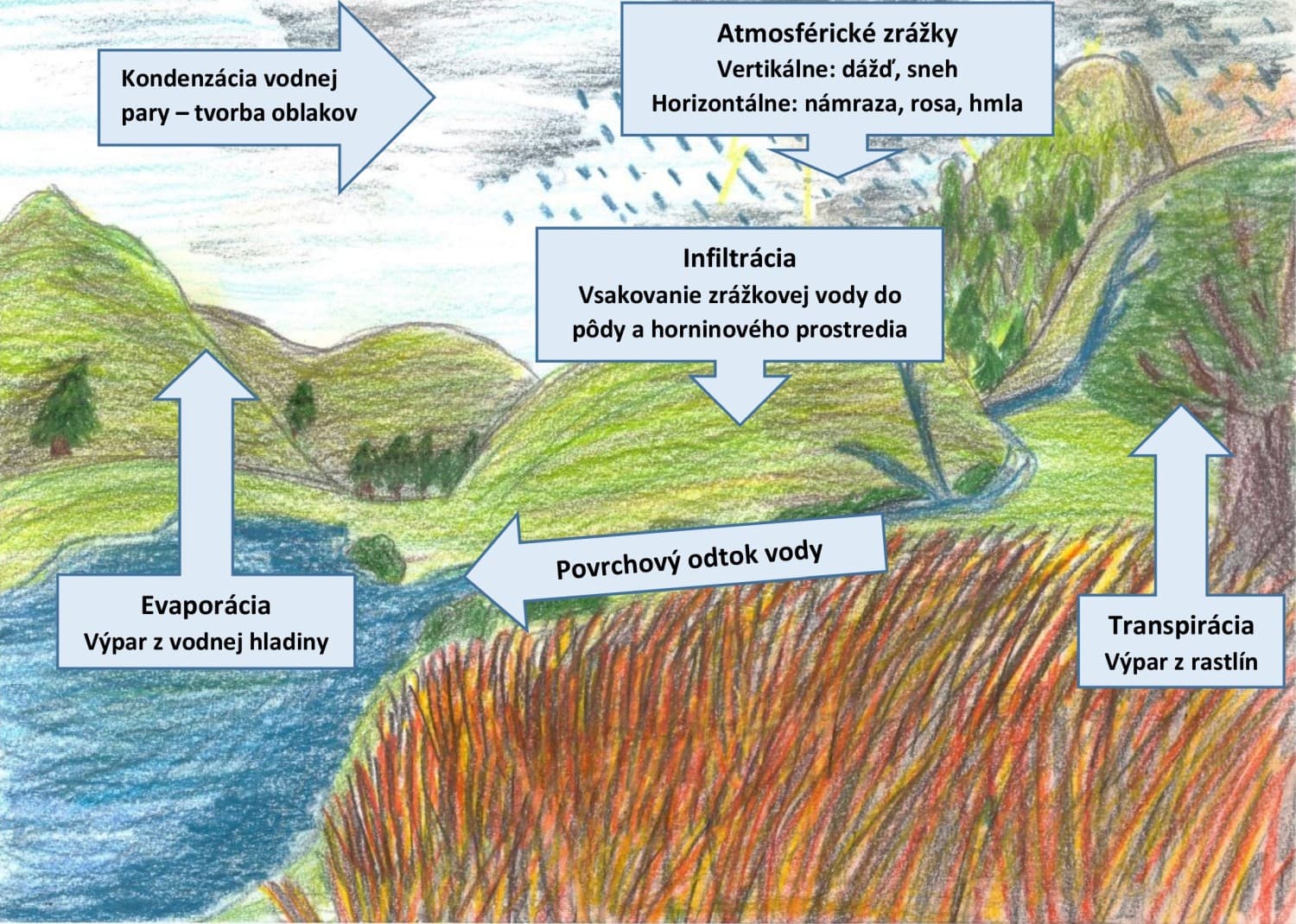
Lokalizácia vody v krajine vytvára špecifický jedinečný ekoton medzi terestrickými a vodnými biotopmi, a preto mu treba venovať náležitú pozornosť, hlavne z pohľadu druhovej aj štrukturálnej diverzity. Vegetácia litorálnych spoločenstiev zahŕňa širokú škálu rastlinných spoločenstiev, kde najdôležitejším faktorom prostredia ovplyňujúcim ich vznik, vývoj a prežívanie je voda. Uplatňuje sa tu predovšetkým povrchová voda, ktorá v závislosti na dobe stagnácie nad povrchom pôdy, výške vodného stĺpca, režimu fluktuácie počas roka zásadným spôsobom zodpovedá za existenciu príslušného vegetačného typu. Vodný režim je prevažne určujúcim faktorom pre spomínanú vegetáciu, ale nie je jediným, ktorý ovplyvňuje formovanie rôznych typov vegetácie mokradí (Ujházy et al. 2013). Vo všeobecnosti môžeme tieto faktory rozdeliť do nasledovných skupín:
- i) klimatické
- ii) hydrologické
- iii) geomorfologické
- iv) biologické interakcie
- v) antropické vplyvy
Medzi klimatické faktory (i) patria najmä: a) teplotný režim vody a substrátu, ale aj vzduchu; teplotný režim vodného prostredia je vyrovnanejší ako v atmosfére vďaka veľkej tepelnej kapacite vody. V sladkovodných jazerách je časť vody na povrchu relatívne teplá a v dolnej časti výraznejšie chladná, a obe vrstvy sú bez výrazného teplotného gradientu, zatiaľ čo medzi nimi figuruje tenšia vrstva s výraznejším kolísaním teploty. Teplota vody zásadným spôsobom ovplyvňuje distribúciu rastlín, ale aj ich fyziológiu, klíčenie, vývojové fázy počas vegetačnej sezóny či prežívanie semien.
Hydrologické charakteristiky (ii), akými sú prúdenie vody, jej hĺbka, krátkodobá či medziročná dynamika, či konektivita sú dôležitými faktormi najmä pre vodnú vegetáciu; okrajovo však ovplyvňujú aj všeobecne vegetáciu mokradí v širšom slova zmysle. Prúdenie vody zásadným spôsobom modifikuje druhové zloženie rastlín na stanovišti; prúdenie zamedzuje prítomnosti voľne vo vode plávajúcich, nezakorenených druhov. Ale aj v stojatej vode má jej pohyb dôležitú úlohu; napr. vlnobitie formuje ako vodnú, tak litorálnu vegetáciu. Hĺbka vody, ako aj environmentálne charakteristiky priamo či nepriamo s ňou súvisiace (napr. teplotný režim, priehľadnosť) rovnako formujú druhovú skladbu a zonáciu rastlinných spoločenstiev. Jedným z najdôležitejších hydrologických charakteristík je kolísanie vodného stĺpca, či už počas sezónny alebo medziročne. Niektoré druhy či ich porasty sú viazané na relatívne ustálený režim s minimálnymi zmenami vo výške hladiny vody, zatiaľ čo iné vyžadujú rozkolísaný vodný režim sprevádzaný jarnými záplavami a letným poklesom vody pod úroveň povrchu pôdy. Hydrologická konektivita je prepojenie rôznych typov vodných biotopov navzájom. Rozlišujeme tak viaceré skupiny konektivity napr. vodné toky (rieky, potoky), biotopy so stojatou vodou (ramená, nádrže, materiálové jamy) bez alebo s trvalým či sezónnym prepojením s vodnými tokmi.
Geomorfologické faktory (iii) ako morfológia či geológia rovnako významne ovplyvňujú existenciu a formovanie vegetácie mokradí. Či už priamo alebo nepriamo vplývajú na svetelný režim (tento je tiež ovplyvnený biologickými zložkami, akými sú napr. pobrežná vegetácia či priehľadnosť vody), charakteristiky substrátu, obsah a kolobeh živín vo vode a v pôde, ale aj chemizmus vody a substrátu. Rovnako je dôležitá veľkosť biotopu; pri terestrických spoločenstvách počet druhov s nárastom plochy rastie, zatiaľ čo pri vodných biotopoch nie je tento trend vždy zrejmý, čo súvisí so špecifickými podmienkami prostredia týchto spoločenstiev.
Svetelný režim je kritickým faktorom pre fotosyntézu, ktorá je nevyhnutná pre život podstatnej väčšiny vyšších rastlín. Je známe, že ak vo vodnom stĺpci klesne svetelná intenzita pod 1 % intenzity dopadajúcej na vodnú hladinu, nie je možná existencia fotosyntetizujúcich vodných rastlín. Štruktúra substrátu môže byť rovnako veľmi dôležitým faktorom determinujúcim prítomnosť rôznych druhov či ich spoločenstiev. Základné delenie podľa veľkosti substrátu je na jemnozrnný organický (napr. bahno, sapropel) alebo anorganický (piesok) materiál, štrk, kamene a balvany, prípadne ich antropické ekvivalenty (napr. dlaždice). Koreniace rastliny majú rôzne stratégie akým spôsobom osídlujú rôzne typy substrátu. Hrubozrnný substrát, kamene a balvany sú prijateľné len pre úzku skupinu vodných makrofytov. Veľká časť vodných a močiarnych vyšších rastlín nachádza optimálne podmienky v jemnozrnejších sedimentoch. Naopak, väčšina machorastov nie je schopná rásť v jemnozrnnom organickom substráte. Dnový substrát zároveň poskytuje pre koreniace rastliny potrebné živiny. Obsah pre rastliny dostupných živín je jedným zo základných faktorov ovplyvňujúcich ich životný cyklus. Podľa ich obsahu vo všeobecnosti rozlišujeme: a) eutrofné vody, bohaté na minerálne živiny s intenzívnym rozvojom planktónu, vyskytujúce sa najmä v teplých oblastiach, b) oligotrofné vody s nízkou koncentráciou minerálnych živín, nižšou hustotou a produkciou planktónu, vyskytujúce sa najmä v horských polohách alebo v oblastiach s oceánickou klímou a minerálne chudobným podložím a c) dystrofné vody, chudobné na minerálne živiny i vápnik, s vysokým obsahom koloidných humusových látok a kyslou reakciou, vyskytujúce sa predovšetkým na vrchoviskách. Ďalšou veľkou skupinou faktorov sú chemické charakteristiky prostredia, akými sú reakcia (pH), alkalinita, salinita (obsah rozpustných solí), vodivosť (obsah rozpustných minerálych látok) vody a pôdy, ako aj obsah rozpustného kyslíka alebo niektorých chemických prvkov v nich.
Biologické interakcie (iv) zahŕňajú biologické charakteristiky jednotlivých druhov, kompetičné vzťahy medzi rastlinami navzájom, prítomnosť herbivorných (rastlinnožravých) druhov živočíchov, alebo choroby rastlín. Na základe biologických vlastností druhov je možné ich delnie na viaceré skupiny, napr. ruderálne tzv. r-stratégovia (napr. Sparganium emersum), druhy ktoré dokážu prežívať v podmienkach s častými/trvalými disturbačnými procesmi (disturbancia – narušenie ekosystému), strestolerantné, dobre reagujúce na nepravidelné disturbačné procesy, konkurenčne silné druhy – kompetítory (napr. Myriophyllum spicatum, Typha latifolia).
Poslednou skupinou faktorov sú vplyvy človeka (v). Nárastom ľudskej populácie, využívaním prírodných zdrojov a krajiny, prudko rastie ich vplyv. Radíme sem ako priame (napr. odvodňovanie mokradí, regulácia a znečisťovanie tokov, likvidácia mokraďových ekosystémov), tak aj nepriame (napr. intenzívne poľnohospodárske využívanie krajiny v okolí mokradí, prísun živín vplyvom hnojenia, skládky odpadov) vplyvy ľudskej činnosti. Pre túto skupinu sa používa aj termín antropogénne faktory.
Význam brehových porastov z hľadiska diverzity
Ripariálna zóna vo všeobecnosti plní sedem základných funkcií (Hansen et al. 2010), ktoré sú dôležité z hľadiska udržateľného rozvoja, a ktoré je potrebné poznávať, manažovať a monitorovať:
- 1) zlepšuje kvalitu vody vo vodnom toku, t. j. redukuje nadbytočný prísun živín a kontaminantov z okolia (nivy, svahov, ciest, polí a pod.),
- 2) redukuje eróziu brehu a prísun sedimentov do koryta,
- 3) zvyšuje zatienenosť koryta, a tým reguluje teplotu vody v koryte,
- 4) zabezpečuje prísun drevnej hmoty, lístia a iných pre akvatický systém dôležitých zdrojov do koryta, t. j. napomáha transferu zdrojov medzi terestrickým a vodným prostredím,
- 5) zvyšuje biodiverzitu koryta,
- 6) autoreguluje a ekologicky prirodzene optimalizuje štruktúru a zloženie svojich rastlinných a živočíšnych spoločenstiev,
- 7) zabezpečuje laterálnu, longitudinálnu a vertikálnu konektivitu riečnej krajiny, slúži ako biokoridor a prostredie transferu podzemnej vody.
K uvedeným funkciám môžeme doplniť ešte ďalšie, ktoré sú z hľadiska udržateľného rozvoja ripariálnej zóny takisto dôležité, a to:
- 8) zmierňuje výšku a rýchlosť povodňových vĺn,
- 9) svojou drsnosťou a vertikálnou akréciou diferencovane akumuluje a filtruje povodňové sedimenty a podmieňuje vývoj fluvizemí,
- 10) generuje prírodný kapitál vo forme zdrojov drevnej hmoty a rastlinných a živočíšnych druhov,
- 11) svojimi estetickými hodnotami dotvára celkový ráz riečnej krajiny,
- 12) poskytuje priestor pre rekreačné, športové a ozdravovacie aktivity,
- 13) slúži ako vhodný objekt pre edukáciu.
Tiež plnia aj funkciu ochrany pred vodou pritekajúcou zo svahov a spôsobujú pokles množstva prívalovej vody a stabilizujú brehovú líniu. Ochranu pred zanášaním a zarastaním plnia vďaka zatieneniu koryta –znižujú prehriatie vody a znižujú prístup svetla. Táto ich funkcia je dôležitá hlavne vo vegetačnom období a pri nízkych vodných stavoch, kedy sa v nezatienených úsekoch rozširujú vodné rastliny (na dne a na svahoch). Tie zvyšujú stupeň drsnosti koryta a znižujú prietokovú plochu. So zvýšeným stupňom drsnosti súvisí aj zníženie rýchlosti, čo má za následok usadzovanie jemných splavenín a plavenín, ktoré postupne zanášajú koryto. Toto nebezpečie sa prejavuje hlavne pri malých tokoch s malým sklonom dna, kde sú následne potrebné časté zásahy. Zatienené koryto spôsobuje teda ochladzovanie vody, zníženie zarastania dna, zníženie jeho zanášania, pričom prípadné vzniknuté nánosy nie sú spevnené vodným rastlinstvom a pri veľkých prietokoch sú odnášané. Zlepšenie samočistiacej schopnosti toku. Táto funkcia je v čiastočnom rozpore s predchádzajúcou funkciou, pretože pre samočistiacu schopnosť vody je potrebný čiastočný prístup svetla.
Samočistiaca schopnosť vody je vlastnosť vody, kedy v tejto nastáva rozklad organických látok až na látky anorganické. Prebieha v závislosti od prítomnosti živých organizmov vo vode, od prítomnosti kyslíka, od teploty a od pohybu vody. Organické látky rozkladajú organizmy nazývané deštruenti. Produkty ich látkovej výmeny konzumuje skupina organizmov nazývaná konzumenti. Samočistenie vody prebieha na rastlinách, na ponorených častiach koreňov, na vetvách a rôznych predmetoch, všade tam kde sa usadzujú kolónie polysapróbnych baktérií rozkladajúcich organické látky. Estetickou funkciou je ich účasť pri spoluvytváraní krajiny.
Umelé úpravy tokov pôsobia ako nevhodný a neestetický prvok. Brehové porasty krajinu naopak harmonizujú vytváraním líniových porastov lemujúcich toky. V prípade nevykonania výsadby po úprave sa brehy oživia náletmi stromov, ktoré nie vždy majú optimálne zloženie a môžu spôsobiť nevhodnú stabilizáciu brehov.
Zoocenologická funkcia spočíva vo vytváraní existenčných podmienok pre tvorbu úkrytov a zdrojov potravy živočíchov. Vytvára vhodné priestory pre rozmnožovanie. Brehové porasty vytvárajú vhodné podmienky pre život rôznych opeľovačov rastlín, hmyzích parazitov a predátorov. Väčšina druhov stromov a krov v brehových porastoch kvitne skoro na jar a v tomto období predstavuje významný zdroj potravy pre opeľovače. Nevýhody brehových porastov sú v:
- • zábere poľnohospodárskej pôdy,
- • neprístupnosti k toku,
- • zhoršení údržby a čistení koryta mechanizmami,
- • zatienení priľahlých pozemkov a opadávaní listov na ich plochu,
- • koreňovej konkurencii a odčerpávaní živín poľnohospodárskym plodinám,- nežiadúcom rozrastaní výmladkov a náletov,
- • zarastaní koreňov do drenáže,
- • tom, že trvalú ochranu tvoria až po niekoľkých rokoch,
- • potrebe pestovných a ťažobných zásahov pri vrastaní do prietočného profilu,
- • možnosti tvorby vývratov,
- • narušenie stability,
- • nebezpečenstvo ľadových zápch pri väčších prietokoch.
Definíciou a hodnotením tzv. funkčných typov a modelov brehových porastov sa podrobne zaoberal VALTÝNI (1981). Citovaný autor uvádza, že funkčný typ brehového porastu je typová jednotka, ktorá predstavuje kombináciu najvýznamnejších funkcií a tým aj funkčné zameranie porastu. Funkčný typ brehového porastu odvodený od funkčných požiadaviek konkrétnej funkcie je reprezentovaný modelom funkčného typu, t. j. modelovou štruktúrou porastu, vrátane zastúpenia drevín, ktoré sa odvíja od ekologických podmienok konkrétnych lokalít. VALTÝNI (1981) uvádza, že z hľadiska ochrany svahov koryta vodného toku pred eróziou je vhodný model brehoochranného funkčného typu brehového porastu. Tento model funkčného typu je možné charakterizovať ako brehový porast s radovo-pruhovým usporiadaním, dvojetážový, s hlúčikovou horizontálnou výstavbou. Prvú (spodnú) etáž majú vytvárať ľahké kroviny s hustými pod povrchovými koreňovými sústavami. Druhú (hornú) etáž majú tvoriť riedke žrďoviny, prípadne kmeňoviny, stromového vzrastu, ktoré majú kolovité alebo srdcovité koreňové sústavy, napr. Fraxinus sp., Quercus sp., Acer sp., Alnus sp. a pod.. Zastúpenie drevín závisí vždy od ekologických podmienok konkrétnych stanovíšť.
Biotopy brehových porastov
Variabilita hydrologických, klimatických a pedologických podmienok viedla k diferenciácií biotopov spojených s povrchovou a sčasti podzemnou vodou. Prehľad biotopov brehových porastov je spracovaný podľa Atlasu biotopov Slovenska.
Ls1 Lužné lesy: Hygrofilné až mezohygrofilné lesy v alúviách rieka potokov (menej popri horských prameniskách), zväčša s pravidelnými alebo občasnými povrchovými záplavami alebo podmáčané podzemnou vodou. Patria sem:
Ls1.1 Vŕbovo-topoľové nížinné lužné lesy: Vŕbovo-topoľové porasty (mäkký lužný les) v najnižších miestach údolných nív väčších riek, na nivných pôdach bohatých na živiny. Hlavným ekologickým faktorom sú pravidelné záplavy povrchovou vodou. Porasty nie sú úplne zapojené, sú spravidla viacposchodové. Krovinové poschodie je druhovo chudobné, prevládajú v ňom zmladené jedince stromov. V bylinnej vrstve sa uplatňujú hygrofilné a nitrofilnédruhy. Typickým znakom je vysoká pokryvnosť a prevaha niektorých rýchlo sa šíriacich autochtónnych druhov, napr. Urtica dioica, Phalaroides arundinacea, Rubus caesius, ale aj zavlečených inváznych druhov, ako sú Aster sp., Solidago canadensis, S. gigantea, Impatiens glandulifera a iné.
Ls1.2 Dubovo-brestovo-jaseňové nížinné lužné lesy: Dubovo-brestovo-jaseňové lužné lesy (tvrdý lužný les) na vyšších a relatívne suchších stanovištiach údolných nív so zriedkavejšími a časovo kratšími povrchovými záplavami. Pôdy sú od typologicky nevyvinutých nivných a glejových až po hnedé pôdy bohaté na živiny. Krovinové poschodie je dobre vyvinuté a druhovo bohaté, v bylinnej vrstve sú prítomné nitrofilné, mezofilné a hygrofilné druhy s výrazným jarným aspektom.
Ls1.3 Jaseňovo-jelšové podhorské lužné lesy: Jaseňovo-jelšové lesy v užších údolných nivách potokov a menších riek ovplyvňovaných povrchovými záplavami alebo podmáčaných prúdiacou podzemnou vodou. Menej typickým stanovišťom sú svahové prameniská alebo terénne zníženiny, kde podzemná voda stagnuje blízko pod povrchom pôdy. Pôdy sú hlinité, stredne ťažké, niekedy oglejené, humózne, s dostatkom živín. Porasty sú spravidla viacposchodové, krovinové poschodie je druhovo bohaté. V bylinnej synúzii sa charakteristicky uplatňujú nitrofilné a hygrofilné druhy.
Ls1.4 Horské jelšové lužné lesy: Porasty jelše sivej s prímesou smreka, zriedkavo ďalších drevín na brehoch horských tokov v chladných údoliach. Pôdy sú piesočnaté, štrkovité až kamenisté. Typická je viac poschodová štruktúra, v krovinovom poschodí dominujú zmladené jedince jelše. V bylinnom podraste sa charakteristicky uplatňujú nitrofilné a hygrofilné druhy. V porastoch asociácie Piceo-Alnetum, stojacich na prechode k podmáčaným smrekovým lesom, je na kyslých mokrých až zbahnených glejových pôdach hojnejšia jelša lepkavá. Z bylín sa hojne objavuje druh Calamagrostis villosa.
Kr8 Vŕbové kroviny stojatých vôd: Uzavreté porasty krovitých vŕb, charakteristické bochníkovitým tvarom a sivou monotónnou farbou s dominanciou vŕb (Salix cinerea, S. aurita), dorastajúce do výšky 2 – 5 (7) m. V bylinnom poschodí, ak v porastoch nestagnuje voda, sa vyskytujú hygrofilné až mezické druhy. Druhové zloženie je závislé od vlhkostných, pôdnych a hypsometrických pomerov. Vznikajú rôzne typy týchto krovín s odlišným podrastom, napr. s druhmi Sphagnum sp., Thelypteris palustris,Rubus caesius, Hydrocharis morsus-ranae, ale aj vysokými ostricami a pod.
Kr 9 Vŕbové kroviny na zaplavovaných brehoch riek: Uzavreté, alebo rozvoľnené krovinaté porasty, často bochníkovitého tvaru, žltozelenej alebo sviežozelenej farby s dominanciou vŕb. Lemujú brehy menších i väčších vodných tokov a ich ramien, ojedinele brehy vodných nádrží a rybníkov. Vŕby dorastajú do výšky 2 – 5 (8) m a dopĺňajú ich niektoré liany a lianely. Porasty sú svetlomilné, pri silnejšom zatienení poschodia stromov tieto zložky ustupujú. Bylinné poschodie je v uzavretých porastoch slabo vyvinuté, v rozvoľnenejších je floristicky bohatšie, zložené najmä z vlhkomilných a nitrofilných druhov. Často na biotopy prenikajú druhy splavené z okolitých lesných a lúčnych porastov. Nachádzajú sa na mladých riečnych naplaveninách tvorených kameňmi, štrkom, štrkopieskom a pieskom.
Br1 - Štrkové lavice bez vegetácie: Útvary okolo riek a väčších horských a podhorských potokov, ktoré na určitých úsekoch ukladajú štrk a hrubší piesok vo forme štrkových lavíc. Následkom rozkolísaného vodného režimu sa na substráte uchytávajú len v minimálnej miere rastliny, zväčša ide o jednoročné a rýchlo klíčiace dvojročné rastliny (Barbarea vulgaris, Filaginella uliginosa, Persicaria lapathifo-lia), ktoré však netvoria pravé fytocenózy.
Br2 - Horské vodné toky a bylinné porasty pozdĺž ich brehov: Trávnaté, prípadne vysokobylinné dvoj- až trojvrstvové spoločenstvá, druhovo chudobné v dôsledku dominancie druhov Calamagrostis pseudophragmites a Phalaroides arundinacea. Ich stanovišťom sú poriečne náplavy podmáčané a podomieľané prúdiacou vodou, kde sa strieda litorálna a terestrická ekofáza. Náplavy sú vzhľadom na rýchlejšie prúdenie hrubozrnnejšie, štrkovité až kamenité. Jemnozem sa akumuluje len medzi kameňmi alebo vytvára na povrchu súvislú vrstvu a tvorí nános v hrúbke niekoľko centimetrov. Porasty tvoria na brehoch tokov charakteristické lemy rôznej dĺžky a šírky.
Br3 - Horské vodné toky a ich drevinová vegetácia s myrikovkou nemeckou (Myricaria germanica): Na území Slovenska osídľuje Myricaria germanica erózne pretvárané štrkové sedimenty na brehoch niektorých horských tokov a tokov vo flyšovej oblasti. Vytvára bylinné i krovinové porasty, v ktorých dominuje. Iniciálne bylinné porasty s Myricaria germanica, ktoré sa môžu mapovať aj ako pod jednotka Br3a, sprevádzajú druhy Acetosa scutata, Calamagrostis pseudophragmites, Chamerion dodonaei, Tussilago farfara, ako aj viaceré náhodne splavené rastliny. V prípade, že sú porasty krovinové a staršie, sú dvojposchodové, pričom Myricaria germanica v nich drevnatie a dosahuje výšku okolo 2 – 2,5 m.
Br4 - Horské vodné toky a ich drevinová vegetácia s vŕbou sivou (Salix elaeagnos): Pionierske spoločenstvo s vyvinutým 5 – 6 m vysokým krovitým poschodím, v ktorom dominuje Salix elaeagnos a S. purpurea, ktoré sú ojedinele doplnené o Alnus incana, Lonicera nigra, Picea abies, Salix fragilis a i. V pestrom bylinnom poschodí sú okrem hygrofilných a subhygrofilných druhov prítomné aj druhy vodou splavené z okolitých lesných a prameniskových spoločenstiev. Porasty lemujú v úzkych pásoch horské bystriny s rýchlo prúdiacou vodou v úzkych dolinách na štrkových, kamenitých, zriedkavo piesočnatých pôdach. Nevyskytujú sa v otvorenej poľnohospodárskej krajine.
Br5 - Rieky s bahnitými až piesočnatými brehmi s vegetáciou zväzov Chenopodion rubri p.p. a Bidention p.p.: Jedno- až dvojvrstvové prirodzené mezotrofné, terofytné spoločenstvá s neskoroletným optimom vývoja v druhej polovici vegetačného obdobia, ale aj spoločenstvá plazivých hemikryptofytov s jarným vývojom. Vyvíjajú sa na obnažených bahnitých a piesočnatých brehoch tečúcich vôd najmä v zátokách, kde pôsobí spätný tlak, alebo na miestach vzdialenejších od riečiska, kde nie je silný prúd vody. V závislosti od dĺžky obnaženia brehov sa nemusia vyvíjať každý rok. Naplavené sedimenty sú pravidelne obohacované živinami, sú rôznej hrúbky (15 a viac cm) a rôznej veľkosti. V dôsledku toho aj porasty kopírujú veľkosť sedimentov, väčšinou sú maloplošné.
Br6 Brehové porasty deväťsilov: Príbrežné spoločenstvá s deväťsilmi (Petasites kablikianus, P. hybridus, menej P. interscendens) alebo Rumex alpinus tvoria fyziognomicky jednotné, husté, zapojené viacvrstvové porasty. Hlavnú vrstvu porastov tvoria rozprestreté čepele listov dominantných druhov vo výške 100 – 160 cm, na živných pôdach aj vyššie. Vyskytujú sa na prirodzených, poloprirodzených až ruderalizovaných stanovištiach na brehoch vodných tokov v horských oblastiach, menej na podsvahových prameniskách a v zamokrených porastoch nivných lúk a v priekopách popri cestách. V druhovo pozmenených a ruderalizovaných formách zostupujú pozdĺž vodných tokov do pahorkatín, výnimočne až do nížin.
Br7 - Bylinné lemové spoločenstvá nížinných riek: Vysokobylinné spoločenstvá na brehoch väčších vodných tokov. Fyziognomicky sú veľmi nejednotné, obvykle viacvrstvové s optimom výskytu na brehoch väčších riek v nížinách a pahorkatinách. Nejednotnosť vzhľadu vyplýva zo striedania sa viacerých dominánt. Typické je zastúpenie väčšieho počtu lian a lianel. Porasty sa vyvíjajú na opakovane záplavami narušovaných, ale živinami dobre zásobených brehových stanovištiach. Sú schopné pomerne rýchlo obsadiť obnažené brehy. Narušovanie stanovíšť, líniový charakter porastov a vysoký vplyv ľudských sídiel a komunikácií, ktoré sú lokalizované na brehoch vodných tokov, spôsobujú čiastočné alebo úplné nahradenie týchto spoločenstiev monodominantnými porastmi poriečnych neofytov.
Br8 Bylinné brehové porasty tečúcich vôd: Spoločenstvá s monodominanciou tráv rodu Glyceria, Leersia a Catabrosa. Sprievodné druhy sú zo skupiny hygrofytov s plazivými a zakoreňujúcimi podzemkami. Biotopom sú prevažne nánosy v zátočinách a v meandroch potokov a menších riek na miestach s nižším prietokom vody. Vyžadujú trvalo zamokrené stanovištia. Nánosy pôdy sú piesčité, piesčito hlinité až hlinité s obsahom organických častíc. Preplavovanie substrátu a stály prísun živín sú predpokladom optimálneho vývoja spoločenstiev tohto zväzu. Kontaktnými biotopmi sú porasty zväzu Oenanthion aquaticae, mapovacia jednotka Vo8. Tvoria sa tiež prechody k biotopom bahnitých brehov alebo trstinových porastov zväzu Phragmition communis. Na spoločenstvá zväzu sa viažu niektoré druhy zo skupiny ohrozených a vzácnych taxónov, napr. Apium repens, Calla palustris, Carex hordeistichos, Nasturtium officinale a ďalšie.